Influence of eating habits and habitats on omega-3 composition and growth in farmed fish
Freshwater and herbivorous fish growth is less affected by low PUFA dietary levels

Omega-3 long-chain polyunsaturated fatty acids (n-3 LC-PUFAs) such as eicosapentaenoic acid (20:5n-3, EPA) and docosahexaenoic acid (22:6n-3, DHA) offer protective benefits against various pathological conditions, including atherosclerosis, obesity, inflammation and autoimmune diseases. Marine fish and seafood are the primary sources of omega-3s in the human diet. However, the inclusion of fish oil in aquafeeds is declining due to limited availability, fluctuating prices, and sustainability concerns.
Due to limited availability, fluctuating prices, and sustainability concerns, the aquaculture sector is increasingly replacing fish oil with vegetable oil in aquafeeds. To comprehensively explore the dependence of growth performance and n-3 LC-PUFA composition on dietary n-3 LC-PUFA levels in cultured fish with varying feeding habits (herbivorous, omnivorous, and carnivorous) and habitats (marine and freshwater), we employed a Bayesian meta-analysis (a type of statistical inference that views probabilities as the likelihood of an event happening) to quantitatively analyze data from 81 selected studies from the published literature.
This article – summarized from the original publication (Wu, Y. et al. 2024. Bayesian Meta-Analysis: Impacts of Eating Habits and Habitats on Omega-3 Long-Chain Polyunsaturated Fatty Acid Composition and Growth in Cultured Fish. Animals 2024, 14(14), 2118) – reports on a study that for the first time used a Bayesian meta-analysis approach, collecting research data from 81 published scientific articles to quantitatively analyze the effects of dietary n-3 LC-PUFA levels on their composition and growth performance in cultured fish.
For detailed information on the study design, data collection and analysis, refer to the original publication.

Composition of dietary fatty acids
In the research reports covered, the overall fatty acid percentages of EPA and DHA in feeds were skewed. Therefore, we categorized the different feed treatments in each study into control and experimental groups. The control group comprised the group with the highest percentage of n-3 LC-PUFAs (typically using fish oil as the fat source). The experimental groups were further divided based on the percentage of n-3 LC-PUFAs relative to the control group into four intervals: 0–25 percent, 25–50 percent, 50–75 percent, and 75–100 percent, denoted as 0 < replacement levels, RL ≤ 25, 25 < RL ≤ 50, 50 < RL ≤ 75, and 75 < RL ≤ 100, respectively.
We collected data and, where necessary, calculated the percentages of SFAs, MUFAs, n-3, n-6, as well as EPA and DHA relative to the total fatty acids for the five aforementioned groups. With the increasing replacement of fish oil with vegetable oil, the proportion of EPA and DHA in feeds and the differences in this proportion among studies are gradually decreasing.
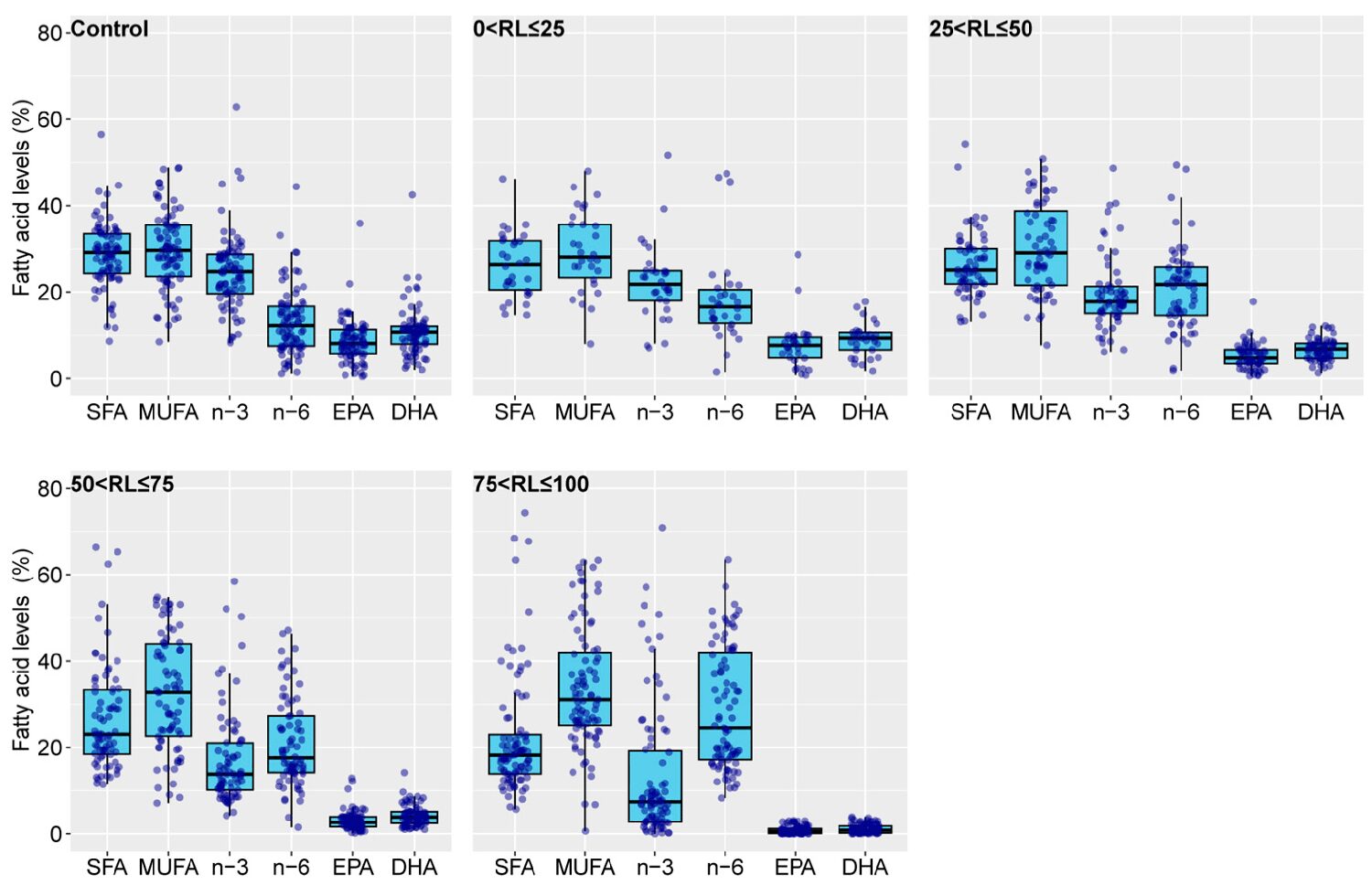
The control group of some research reports showed a proportion of EPA close to zero. These studies administered DHA alone instead of fish oil. The differences between n-3 PUFAs and n-6 PUFAs among groups are consistent with the fact that plant oils generally have lower levels of n-3 PUFAs and a higher content of n-6 PUFAs. Furthermore, the absence of EPA and DHA in plant oils allows for the content of n-3 LC-PUFAs in feeds to directly reflect the level of fish oil substitution by vegetable oil.
Final body weight
A slight reduction in the n-3 LC-PUFA content had a limited impact on the final body weight, FBW, with carnivorous fish even exhibiting a positive effect, as indicated by a 95 percent CrI falling below 0. Of the total of 24 studies analyzed, 10 reported effect-size values within [0.006, 1.379], while the remaining 14 studies had a range within [−0.3061, −8.9756]. Marine fish dominated this classification with 20 studies, while 4 studies focused on freshwater fish, aligning with the distribution characteristics observed in pooled effect-size intervals.
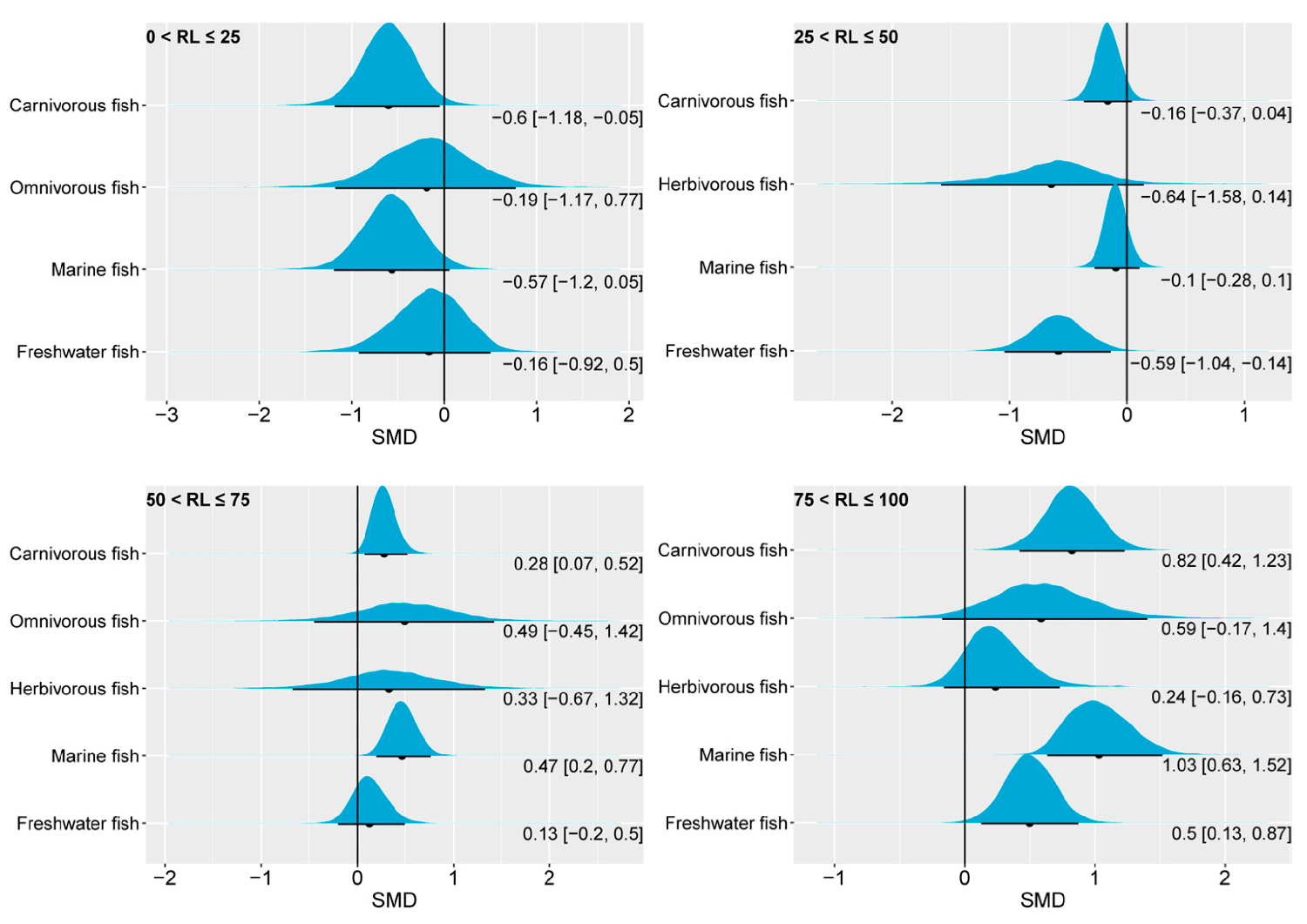
Overall, from the perspective of FBW, our analysis results revealed that marine fish and carnivorous fish have a high demand for dietary n-3 LC-PUFAs, followed by freshwater fish. The FBWs of omnivorous and herbivorous fish, on the other hand, were less responsive to changes in n-3 LC-PUFAs in feed.
Specific growth rate
The pooled effect size of the specific growth rate, SGR, in carnivorous, omnivorous, herbivorous, marine, and freshwater fish showed similarities with the FBW at different oil substitution levels. Carnivorous fish and marine fish did not show significant differences compared to the control when the dietary n-3 LC-PUFA replacement level was less than 50 percent of the value in the control group. However, when the level exceeded 50 percent of the control group, the SGR performances of carnivorous fish and marine fish were suboptimal. Freshwater fish, on the other hand, only exhibited decreased SGR values when the replacement level surpassed 75 percent of the control group. In the present study, herbivorous and omnivorous fish showed the insensitivity of the SGR to changes in the content of dietary n-3 LC-PUFAs.
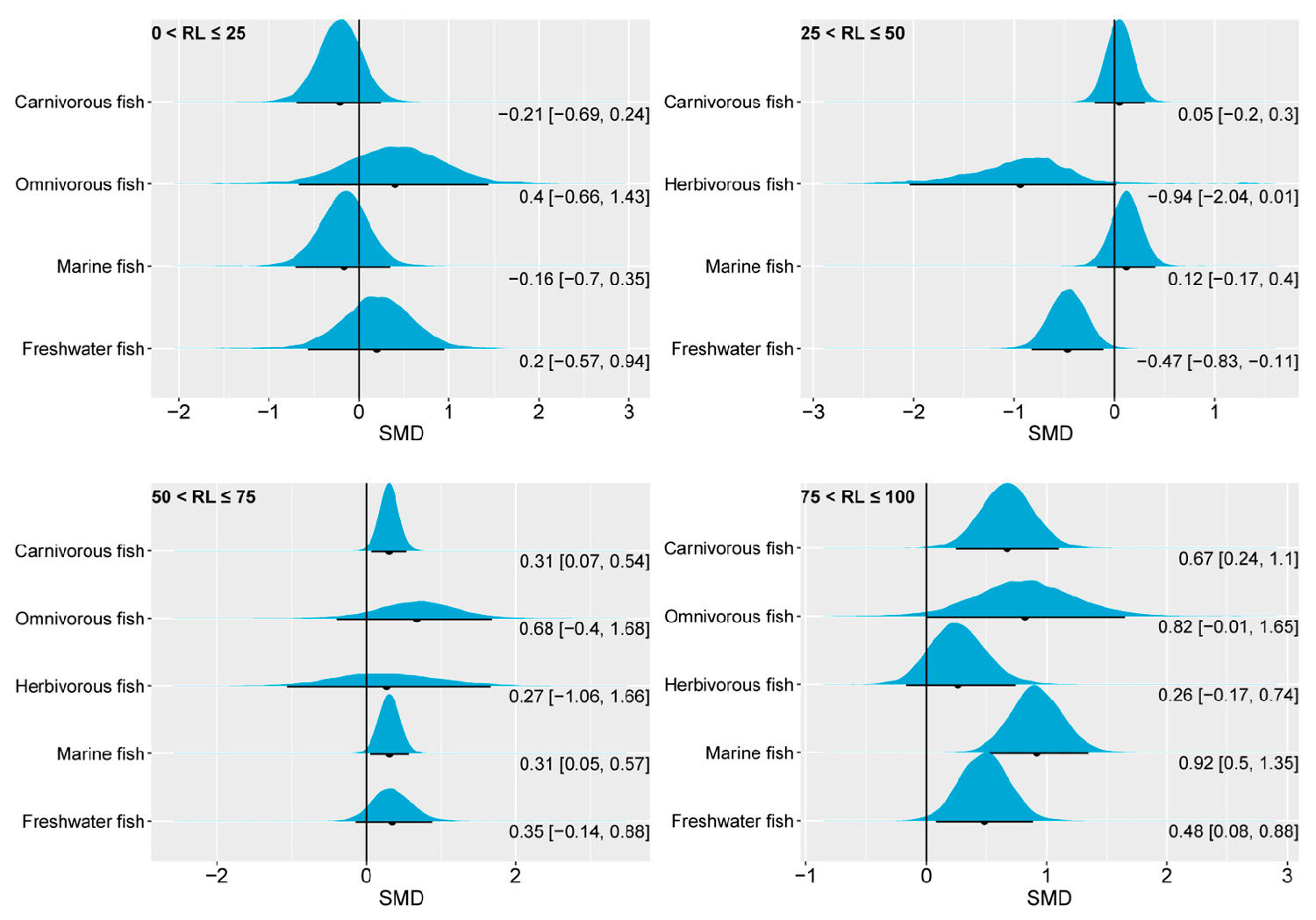
Our findings are consistent with previous studies on growth performance using frequency-based meta-analyses. Other researchers have reported that substituting 50–100 percent of fish oil with various plant oils (including canola oil, linseed oil, and soybean oil) also has an unfavorable impact on fish growth. In addition, the substitution of 100 percent fish oil with plant oil also had a significant negative effect on the growth of both freshwater and marine fish. As in the present study, the impact on marine fish was more pronounced than in freshwater fish. Another study revealed a significantly negative pooled effect size of substituting fish oil with plant oil on the SGR of marine and carnivorous fish. In this regard, there is evidence indicating that dietary plant oil can trigger growth hormone resistance in marine fish. Additionally, the role of dietary n-3 LC-PUFAs in enhancing immune function may collectively contribute to the impact of dietary n-3 LC-PUFA levels on the growth performances of marine and carnivorous fish.
Liver EPA and DHA
The impact of changes in the ratio of dietary n-3 LC-PUFAs on the hepatic content of EPA was generally consistent in different fish species, with higher levels of dietary n-3 LC-PUFAs often resulting in higher liver EPA proportions. This observation is not surprising, as numerous studies indicate that the fatty acid composition in fish body fat broadly reflects the proportion of fatty acids ingested.
We observed differences in the liver EPA content of fish with different feeding habits and habitats at varying substitution levels of n-3 LC-PUFAs in the feed. In carnivorous and marine fish, reducing dietary n-3 LC-PUFAs to 0–25 percent of the control significantly decreased the liver EPA content.
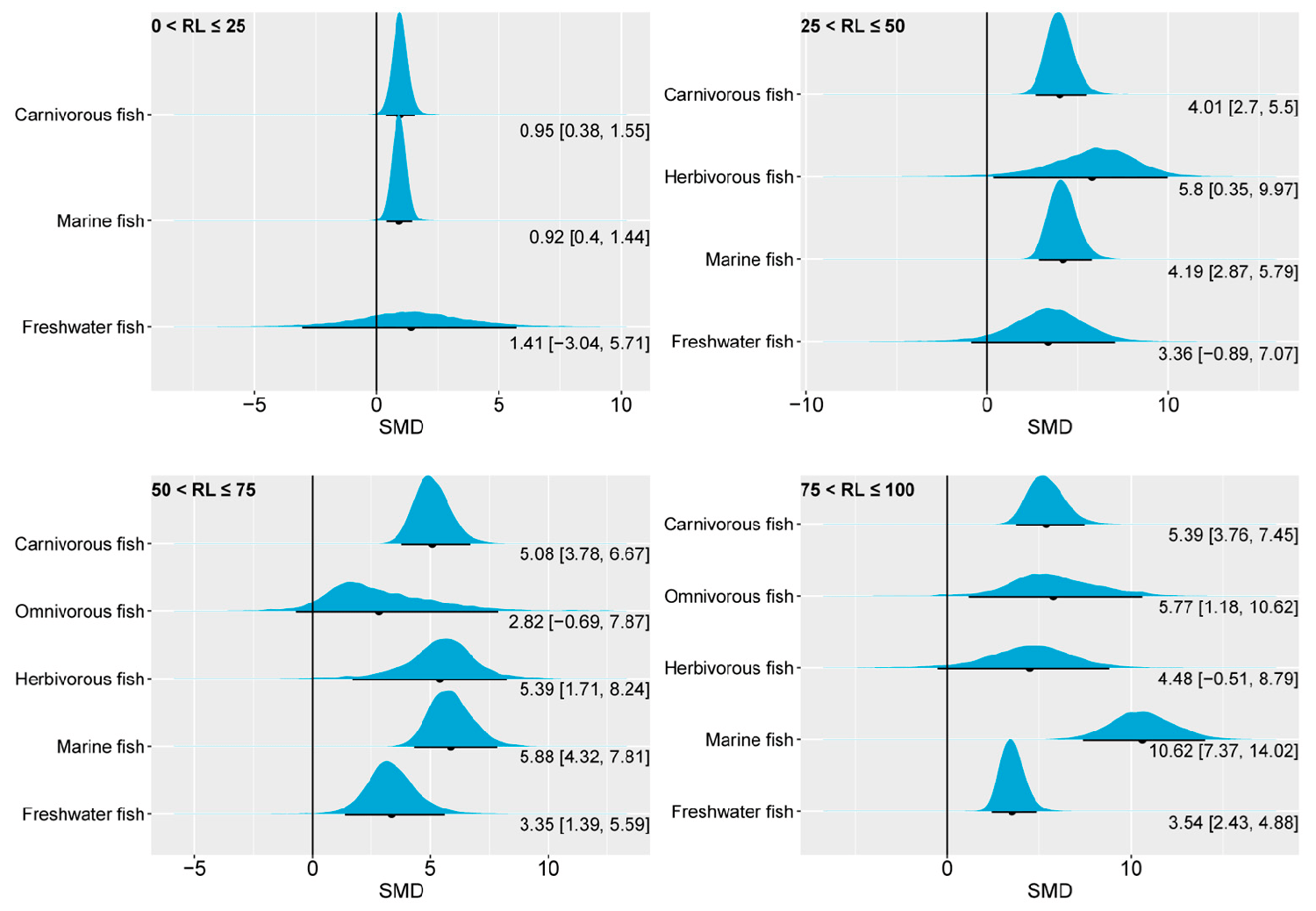
The response of liver DHA to oil substitution and varying dietary n-3 LC-PUFA levels was similar to that of liver EPA, wherein a noticeable decrease in the liver DHA content was observed when aquafeeds contained reduced levels of n-3 LC-PUFAs. When the substitution level was between 0 and 50 percent, there was no significant difference in liver DHA content compared to the control, but when the oil substitution level was between 50 and 100 percent, the 95 percent CrI of liver DHA of freshwater fish showed a clear above-zero range.
For carnivorous fish and marine fish, there was a significant overlap in 95 percent CrI of liver EPA between the 25–50 percent and 50–75 percent oil substitution levels, whereas this phenomenon was less pronounced in the 95 percent CrI of liver DHA. This suggests that liver DHA, compared to EPA, was more dependent on changes in dietary n-3 LC-PUFAs.
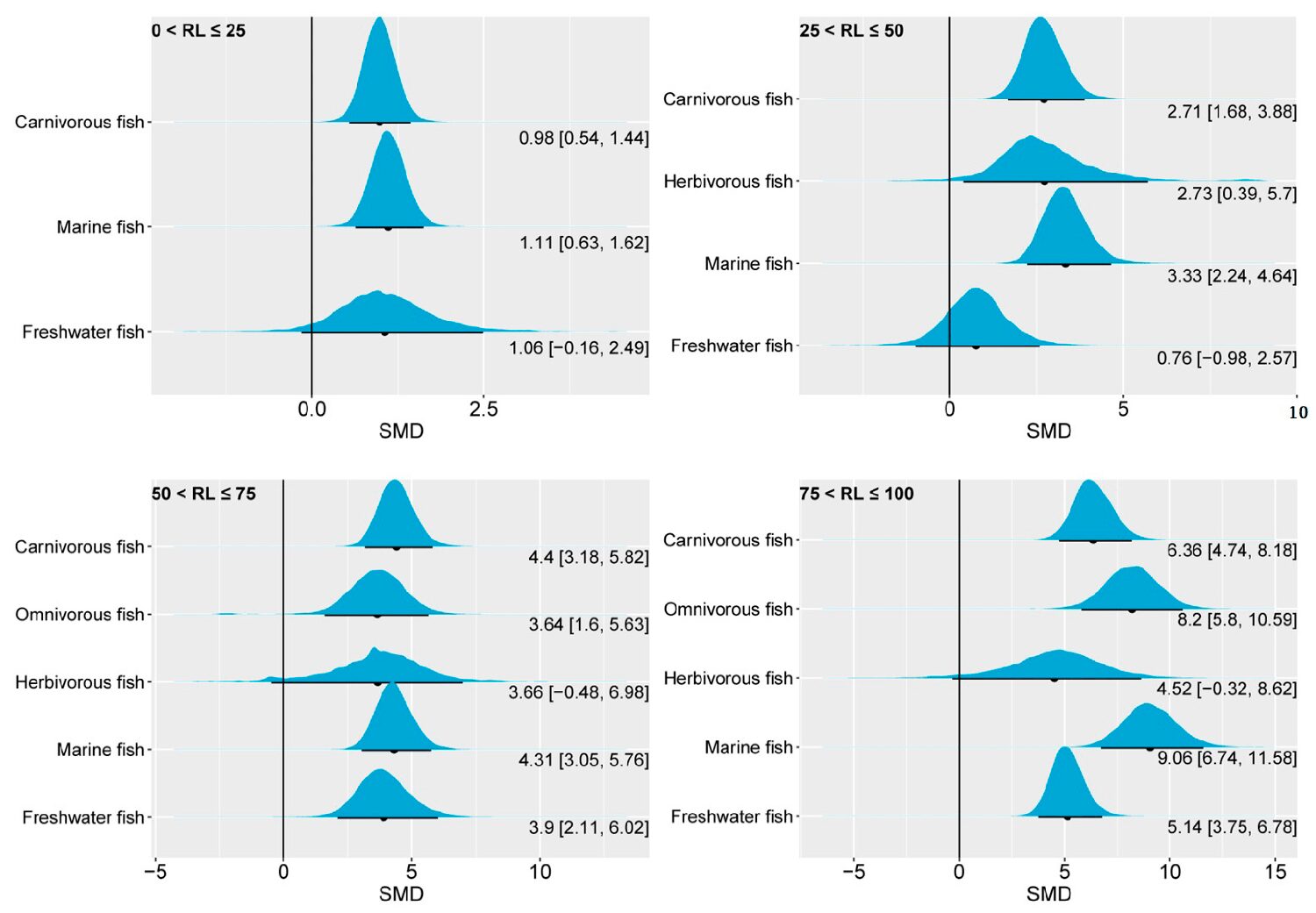
Muscle EPA and DHA
In general, the performance of EPA and DHA in the fish muscle was similar to that in the liver in response to changes in dietary n-3 LC-PUFAs. However, the response of muscle EPA and DHA to changes in dietary n-3 LC-PUFAs appeared to be more sensitive than in the liver. This is consistent with the fact that the skeletal muscle primarily plays a role in retaining EPA and DHA. Nevertheless, the proportions of muscle EPA and DHA exhibited different responses to changes in dietary n-3 LC-PUFAs depending on the fish category.
Regarding muscle EPA, carnivorous and herbivorous fish exhibited remarkable differences in response to changes in dietary n-3 LC-PUFAs. Indeed, in all four groups of this study, the 95 percent CrI of the SMD for muscle EPA in carnivorous fish was significantly greater than 0, indicating that increasing the amount of n-3 LC-PUFAs in the feed can effectively enhance the levels of muscle EPA in this type of fish. On the other hand, the 95 percent CrI of herbivorous fish only demonstrated an above-zero range in the highest oil substitution-level group.
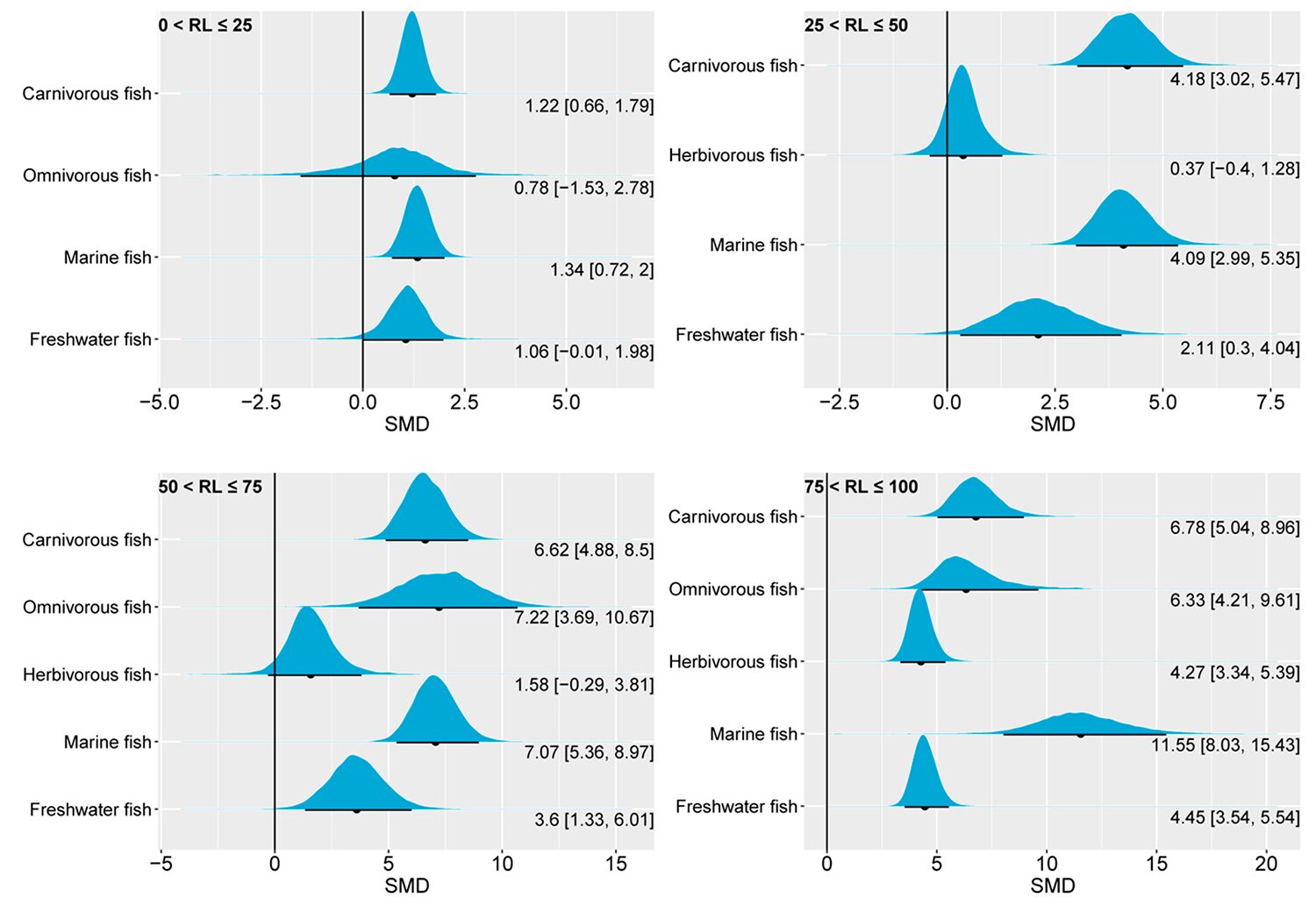
Compared to muscle EPA, muscle DHA showed a higher sensitivity to changes in dietary n-3 LC-PUFA levels. Except for herbivorous fish, the muscle DHA of other fish species in the four groups considered in this study all showed an above-zero range for the 95 percent CrI. Additionally, except for marine fish, which showed a slightly higher 95 percent CrI, the 95 percent CrI of muscle DHA remained similar across fish species as the substitution level increased. Since the biosynthesis of DHA is downstream of EPA, the similarity in the effect of dietary n-3 LC-PUFA levels on the muscle DHA content of different fish species may indicate that their biosynthetic capacities for DHA may be comparable. Since some lower aquatic organisms have the ability to synthesize n-3 LC-PUFAs, the dietary requirements of fish for DHA are likely to meet primarily through the trophic chain.
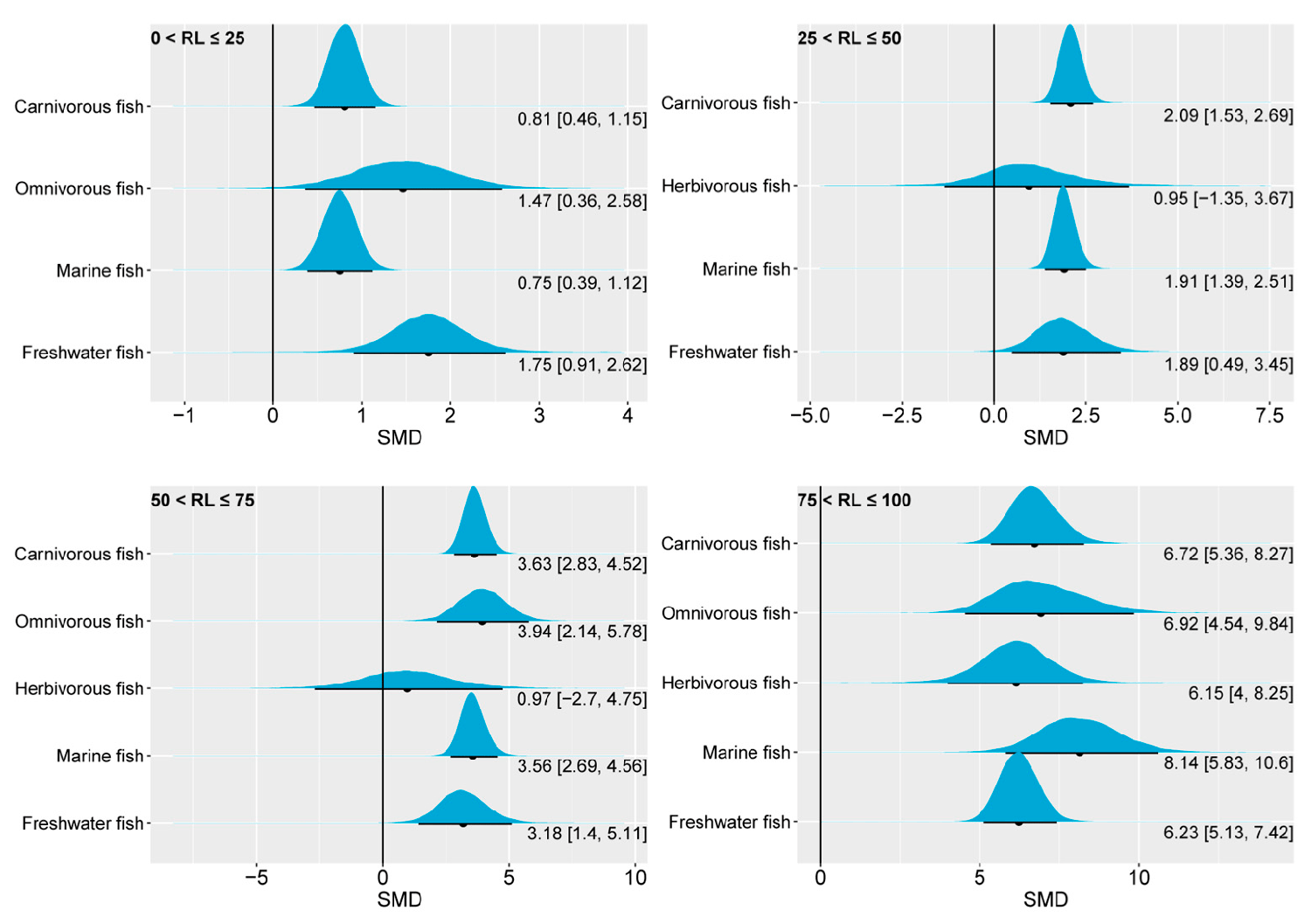
Insights into dietary omega-3 levels affecting growth of marine and carnivorous Fish
The findings of the present study demonstrate that both the growth performance and accumulation of omega-3 fatty acids in marine and carnivorous fish are highly sensitive to the dietary content of n-3 LC-PUFAs. Thus, gaining a deeper understanding of the intermediary metabolic processes involving n-3 LC-PUFAs in marine and carnivorous fish is of significance for improving their aquaculture production and quality.
Unlike terrestrial animals, fish, particularly carnivorous fish, tend to utilize amino acids rather than carbohydrates to supply energy needs. The protein requirement for carnivorous fish is typically around 46 percent. Amino acid usage for energy metabolism in fish is consistent with their weak carbohydrate digestion and breakdown capabilities. Some studies found that fish also tend to utilize amino acid breakdown energy rather than carbohydrates for lipid synthesis.
Although carnivorous fish preferably use amino acids to obtain energy, various studies have demonstrated the potential of carnivorous fish to utilize dietary carbohydrates for growth. A sterol regulatory element-binding protein is a potent transcription factor with a major role in the control of lipid biosynthesis, and also participates in the regulation of carbohydrate metabolism-related enzymes in carnivorous fish.
Modulating the proportion of n-3 LC-PUFAs in feed may be one approach to enhancing the ability of fish, particularly marine and carnivorous species, to utilize dietary carbohydrates. High-carbohydrate or high-fat diets are prone to trigger metabolic dysregulation, and the relatively weak glucose-utilization ability in fish may exacerbate this progression. In teleost fish (about 96 percent of all fish species), dietary supplementation with n-3 LC-PUFAs exerts varying effects on glucose metabolism, but the impact of n-3 LC-PUFAs on resting-state circulatory glucose levels in carnivorous fish appears to be minimal.
The n-3 LC-PUFA-mediated enhancement of fish growth performance, especially in marine and carnivorous species, may be due to the improvement of insulin sensitivity and the consequent utilization of dietary carbohydrates. Our recent report also shows that n-3 LC-PUFAs can improve the growth performances of marine and carnivorous fish, which could be due to improved insulin sensitivity.
However, the impact of dietary n-3 LC-PUFA content on growth performance is not always significant and may interact with other factors such as temperature, dietary lipid content, and salinity. For instance, dietary n-3 LC-PUFAs may not benefit the growth of carnivorous fish at certain replacement levels, and higher water temperatures might enhance the contribution of saturated fatty acids to growth.
Perspectives
This study was the first to employ Bayesian statistical methods and conduct a meta-analysis using omega-3 fatty acid levels in aquafeeds as the primary variable to analyze their impact on n-3 LC-PUFA content and growth in cultured fish. The methodology herein used avoided the impact of residual oil in fishmeal, which is approximately 10 percent w/w and may be an overlooked factor when studying the fish oil requirements of farmed fish.
The results of this study can contribute to the optimization of fish oil levels in aquafeeds and promote the development of more sustainable aquaculture practices. Our findings showed that the growth performances of freshwater and herbivorous fish were less affected by low levels of n-3 LC-PUFAs in the feed. In contrast, for optimal growth of marine fish, dietary n-3 LC-PUFA levels should be at least approximately 7 percent of the total fatty acids (mean value of the 50 < RL ≤ 75 group). In terms of nutritional value, and with the exception of herbivorous fish, changes in the proportion of dietary n-3 LC-PUFAs were directly reflected in the muscle n-3 LC-PUFA content of cultured fish.
Further promising research efforts include incorporating n-3 LC-PUFA-rich algae and bacterial oil into the diet and developing transient gene therapy methods to express exogenous enzymes that enable omega-3 fatty acid biosynthesis in culture fish without producing genetically modified organisms.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Author
-

Corresponding author
Secció de Bioquímica i Biologia Molecular, Departament de Bioquímica i Fisiologia, Facultat de Farmàcia i Ciències de l’Alimentació, Universitat de Barcelona, Joan XXIII 27-31, 08028 Barcelona, Spain
Related Posts

Aquafeeds
Aquaculture Exchange: Lukas Manomaitis, USSEC
The U.S. Soybean Export Council is a huge supporter of aquaculture growth globally, as so many aquafeed formulators rely on U.S. soy to create nutritious diets. The Southeast Asia senior technical advisor for USSEC’s aquaculture program talks about this symbiotic partnership.

Intelligence
Are omega-3s in farmed jade perch as high as believed?
Farmed jade perch – similar to other cultured fish species – is only rich in omega-3 fatty acids if its diet consists of these nutrients in high amounts. How does it compare to the wild fish?

Health & Welfare
Effect of juvenile Atlantic salmon diets with different levels of PUFAs
Study investigates blood and muscle changes in young Atlantic salmon when fed diets containing different amounts of polyunsaturated fatty acids, or PUFAs.

Health & Welfare
Long-chain PUFA requirements of juvenile California yellowtail
This study shows that soybean oil can totally replace fish oil in California yellowtail aquafeeds without affecting production performance and reducing the omega-3 fatty acid profile of fish tissues, as long as acceptable levels of ARA and DHA are provided.