Improving the lipid profile of black soldier fly larvae
An overview of the current status for the manipulation of lipid content of black soldier fly (Hermetia illucens) larvae for marine aquafeeds formulation

The use of insects as aquafeed ingredient and as a potential substitute for fishmeal has significant potential. The natural amino acid profile and the protein content of black soldier fly (BSF, Hermetia illucens) larvae is suited for most aquafeeds and continues to gain growing attention. But regarding its lipid content, the high levels of saturated fatty acids (SFAs) along with the low content of polyunsaturated fatty acids (PUFAs) are risks to the direct use of BSF as a reliable substitute for fish oil when it comes to marine fish or shrimp.
Higher inclusions of insect-based ingredients in the formulation of marine aquafeeds can be an important step towards sustainability. To pursue this goal, the modulation of the lipid profile of BSF larvae has received great attention. While its nutritional profile can shift with diet, the ability to modulate its lipidome [the totality of lipids in cells] is yet to be understood.
This article – adapted and summarized from the original publication [Rodrigues, D.P. et al. 2022. Improving the Lipid Profile of Black Soldier Fly (Hermetia illucens) Larvae for Marine Aquafeeds: Current State of Knowledge. Sustainability 2022, 14(11), 6472] – provides an overview of the lipid modulation of H. illucens larvae through its diet, aiming to produce a more suitable ingredient for marine aquafeeds.
Study setup
To summarize the current trends on the use of different substrates already tested to feed BSF and the achievements concerning the modulation of its lipid fraction, a systematic literature search was carried put, and a total of 148 fatty acid profiles of BSF larvae were reviewed. For detailed information on the literature search; inclusion criteria and data extraction; and statistical analyses, refer to the original publication.

Results and discussion
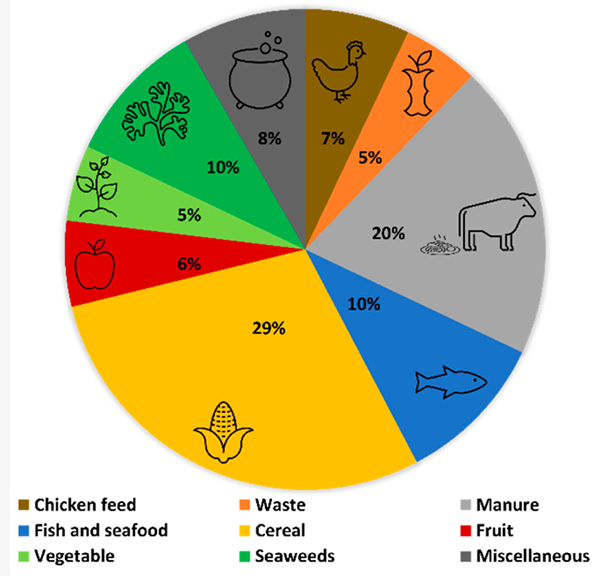
It is well documented that BSF larvae can feed on a multitude of substrates, including manure, fish offal, brewery and winery byproducts, restaurant waste and fruits or vegetables. In this survey, cereal, manure, fish and seafood were the top three substrates used to feed BSF larvae (Fig. 1). These findings highlight that, in recent years, multiple studies have been pursuing the application of BSF larvae as an ingredient in aquafeeds, aiming to enhance its levels of fatty acid unsaturation and long-chain omega-3 PUFAs.
The average lipid profile assembled from our study displays five major contributors for saturated fatty acids (SFAs): lauric acid, palmitic acid, myristic acid, stearic acid, and arachidic acid. Lauric acid is the most abundant fatty acid found in BSF larvae. According to Spranghers, et al., the abundance of this fatty acid is not conditioned by the stage of development of BSF, and when analyzing the data retrieved during our study the present work, this is clearly associated with the feeding substrate provided (Fig. 2). Feed with a marine origin – such as seaweeds, fish, or mussels – present zero or very low values of this fatty acid. The highest values are found in feeding substrates using fruits, vegetables and some cereals.
We demonstrated in our review that several studies already tried, and succeeded, to promote an increase in the PUFA content of BSF larvae, namely on alpha-linolenic acid (ALA), eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). And recent developments show that feeding substrates that produced BSF larvae with the highest contents in omega-3 fatty acids [polyunsaturated fatty acids or PUFAs can be classified in n-3 fatty acids and n-6 fatty acids, and in westernized diets the predominant dietary PUFAs are n-6 fatty acids. Both types of fatty acids are precursors of signaling molecules] were fish discards (39.9 percent), coffee silverskin supplemented with the heterotrophic microalgae Schyzochytrium sp. (28.5 percent) and seaweeds (14.1 percent).
The content of essential fatty acids for marine fish and shrimp (such as EPA and DHA) in BSF larvae was related to the type of substrates used during larval feeding. In the case of ALA, the most significant levels were obtained using chicken feed with a supplementation of 4 percent of flaxseed (9.7 percent) and vegetables (5.8 percent). Relevant levels of EPA and DHA were recorded in BSF larvae fed with fish discards (13.6 percent and 21.4 percent, respectively), as well as when fed with coffee silverskin with Schyzochytrium sp. (11 percent and 16 percent, respectively).
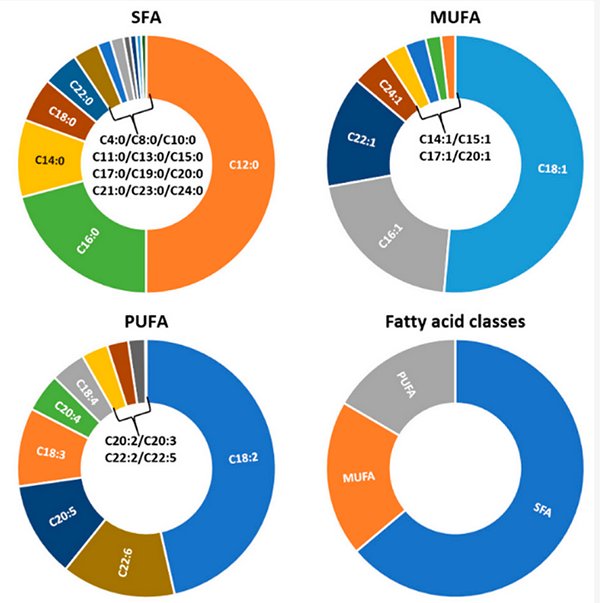
Oleic acid is the most common fatty acid in nature and it was found in all data analyzed in this survey. Results ranged from negligible amounts to a maximum of 32 percent when BSF larvae were provided with cow manure. As part of the fatty acids that constitute fat tissue, palmitoleic acid is widely common and found in most study cases of this review, with the exception of BSF larvae fed with Philippine tung seed, cow manure, cow manure with fish and swine manure. Despite its wide availability, the maximum value registered was 10 percent.
Further analysis of PUFA results show the following fatty acids with a major contribution: linoleic acid, alpha-linoleic acid, DHA, EPA and others. Linoleic acid is one of the most common PUFAs in insects. The n-6 conformation was found in most studies retrieved, with only a few exceptions, such as when feeding BSF larvae with palm kernel meal, ice cream industrial waste, coconut endosperm and soybean crude residue and food waste. The percentages registered ranged from 0.49 percent to 52.5 percent. The n-3 form was found exclusively when using swine manure at 2 percent, dog food, and human feces. Consistently, DHA was detected in BSF larvae reared on marine sources, with fish discards being the substrate that yielded the highest levels of this fatty acid (21.4 percent).
EPA presented the same outline, with marine sources being the privileged source for this fatty acid, again with fish discards as its major contributor (13.6 percent). However, in minor amounts, other substrates like food waste, swine manure, soybean crude residues and others also evidenced this fatty acid in BSF larvae.
Considering alpha-linolenic acid and gamma-linolenic acid, the n-3 form was the dominant conformation detected in most substrates in levels going from 0.19 percent up to 9.7 percent. This fatty acid is of major relevance due to its ability to be bio-converted into highly unsaturated fatty acids by some marine and freshwater fish species. The n-6 conformation was also found, although in lower amounts (0.02 percent to 2.6 percent) and in fewer substrates.
The differences among the fatty acid profiles of larvae supplied different feeding substrates are represented in Fig. 3. These results emphasized the presence of DHA and EPA in seaweeds and seafood and separate fruit and manure as substrates with low levels of ALA, EPA, DHA and PUFA. Furthermore, cereals and fruits presented the lowest levels of SFAs and PUFAs, respectively. The data analysis isolated two major groups, with seaweeds and seafood being separated from all substrates surveyed. Moreover, SFA is separated as well from the other four groups, which were clustered in pairs: DHA with EPA and ALA with PUFA.
![Fig. 3: Heat map [a graphical representation of data where values are depicted by color] of fatty acid profiles of <em>H. illucens</em> larvae fed on different substrates (n = 148) using fatty acid relative abundances (log-transformed and auto-scaled) and ranging from down-accumulated (green) to up-accumulated (red). Adapted from the original.](https://www.globalseafood.org/wp-content/uploads/2022/06/RODRIGUES-Fig-3.png)
However, such substrates do not need to undergo any further processing and may be directly provided to BSF to obtain larvae enriched with n-3 PUFAs and HUFAs. This is not the case when these same marine-based substrates (e.g., fish offal) are used to formulate aquafeeds for marine fish/shrimp. This approach may allow the reduction of production costs of marine aquafeeds, making it possible to incorporate in a single ingredient both balanced protein and lipid contents.
Perspectives
There are clear benefits associated with the mass production of insects using approaches that do not compete with food production for human use, which enhance waste management and nutrient recycling. However, there are still some technological constraints concerning the mass-rearing of insects that need to be overcome. Upscaling strategies allowing continuous production with consistent quantities and quality need to be implemented while reducing production costs and making insect meals more competitive.
Nevertheless, the growing development of mass-rearing systems sets the stage to further explore creative solutions to these current bottlenecks. An in-depth understanding of BSF larval lipid metabolism and lipidome, if supported by trials testing its metabolic plasticity and environmental limits, are key to benefit the most from feeding substrates currently available to mass produce BSF. Moreover, biotechnological approaches should be considered to improve the nutritional bioavailability of substrates and upraise the yields of dietary incorporation, namely for polyunsaturated fatty acids (PUFAs).
Research exploring the potential existence of dormant metabolic pathways on BSF larvae or its associated gut microbiome, that may be triggered to allow the production of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) from alpha-linolenic acid (ALA) precursors must also be pursued. Published results on the modulation of the fatty acid profile of BSF larvae we analyzed are certainly promising but, with a few exceptions, are still unable to fulfill the nutritional needs of marine fish and shrimp species currently being farmed. With a biomass rich in essential amino acids and minerals, it is most certain that in upcoming years, BSF larvae will become a regular ingredient in aquafeeds formulations. Its use will certainly increase for premium batches of this ingredient featuring high levels of essential omega-3 PUFAs and HUFAs.
Our review aims to inspire researchers to pursue innovative approaches that may allow the unleashing of the true potential of BSF as a key ingredient for the formulation of marine aquafeeds, supporting sustainable aquaculture and contributing to the harnessing of blue foods.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Corresponding author
ECOMARE, CESAM–Centre for Environmental and Marine Studies, Department of Biology, University of Aveiro, Santiago University Campus, 3810-193 Aveiro, Portugal -
ECOMARE, CESAM–Centre for Environmental and Marine Studies, Department of Biology, University of Aveiro, Santiago University Campus, 3810-193 Aveiro, Portugal
-
Group of Recycling and Valorisation of Waste Materials (REVAL), Marine Research Institute (IIM-CSIC), C/Eduardo Cabello, 6, 36208 Vigo, Galicia, Spain
-
ECOMARE, CESAM – Centre for Environmental and Marine Studies, Department of Biology, University of Aveiro, Santiago University Campus, 3810-193 Aveiro, Portugal
Related Posts

Aquafeeds
Black soldier fly larval production in a stacked production system
Study describes development and evaluation of an “all-in-one” stacked system for indoor production of black soldier fly larvae.

Health & Welfare
Effect of full-fat, black soldier fly meal in juvenile Siberian sturgeon diets
Assessment of the effects of black soldier fly meal in juvenile Siberian sturgeon diets shows low feed acceptance at high levels.

Aquafeeds
Fly guys: Canada opens the door for insect-based feed companies
The sci-fi flick “The Fly” warned about mixing flies and technology, but high-tech black soldier fly farmers are seizing a real opportunity in aquaculture.

Aquafeeds
Comparing the nutritional value of black soldier fly larvae meal processed with spray- and oven-drying methods
Study from Malaysia shows that the nutritional profiles of black soldier fly larvae meals can vary, depending on the processing methods used.