Domestication and broodstock management
Implications for long-term quality of cultured stocks

A domesticated organism is one that has been trained to live with, or be of service to, humans. Perhaps more practically, it is one whose life cycle has been closed in artificial environments.
Most cultured species today have been domesticated, many since the advent of induced spawning methodologies. However, a significant proportion of aquaculture production, particularly in the Asian shrimp industry, is still derived from wild-caught seed or broodstock.
Since genetic management and improvement can only be applied to stocks whose life cycles are completed in captivity, it is important for those cultured species that remain dependent on wild stocks to be brought under domestication. It is also important to recognize that, as in the introduction of genetically changed populations to indigenous wild stocks, domestication can have potentially negative effects, as well.
Negative impacts
Several studies, including recent research on Nile tilapia and the Indian carp or rohu, have illustrated the negative impacts of domestication by demonstrating the superior performance of wild-caught over their domesticated counterparts. In such cases, it is wise to go back to the wild stocks – which are presumed to have greater genetic variation – as the basis for future stock development. This is indeed what was done in genetic-improvement programs for both of these species.

Domestication processes
Due to new selective pressures in aquaculture environments, the domestication process must be properly conducted. Broodstock management must avoid undesirable changes in gene frequencies through inbreeding, genetic drift, hybrid introgression, and unconscious selection. Unfortunately, many domestication programs – often those carried out on an ad hoc basis using whatever stock may be readily available at the time – have not paid due regard to these processes.
Founder effects
Founder effects, modified by subsequent broodstock management practices, have brought about major changes in gene frequencies and consequent measurable variations in the culture potential of many domesticated aquatic organisms. A dramatic example is the introduction and subsequent spread of the tilapia (Oreochromis mossambicus) throughout Asia and the rest of the world.
Five specimens of the species, indigenous to southeastern Africa, were first discovered in Indonesia in the late 1930s. These were bred and, in the absence of any information on subsequent fresh introductions from Africa, are assumed to be the ancestors of all cultured and feral populations of the species throughout Asia.
Severe genetic bottleneck
A recent study of several feral populations of Oreochromis mossambicus in Asia revealed they had a sixfold reduction in allelic diversity and up to threefold reduction in heterozygosity levels when compared to wild-caught stocks in southern Africa. Founder effects due to this severe genetic bottleneck early in the process of domestication have thus had profound impact on the levels of genetic variation.
Limited culture potential
Furthermore, the culture potential of these Asian populations appears very limited, to the extent that they have been largely displaced in aquaculture by O. niloticus. It is likely that significant loss of genetic variation has occurred in most domesticated aquaculture stocks. The major factor in avoiding the most negative consequences of initial domestication is to ensure the genetic contribution from the maximum number of individual founding stocks.
Local adaptation

The negative impacts of reduced genetic variation in domesticated stocks are often improved by the positive effects of local adaptation from unconscious selection. There is very strong selection pressure against genotypes (the particular set of genes possessed by the individual fish) that cannot adapt to the artificial environment – particularly during the early phase of domestication. Domesticated stocks are therefore intensively selected for adaptation.
A case in point is the indigenous common carp used in traditional rice-fish culture systems in northern Vietnam. These carp stocks, which have been maintained as reproductively isolated populations within the rice-fish farm communities, are likely to be highly inbred – a major factor behind their poor growth rates.
However, the fish are well adapted to the often extreme environment of the rice fields, particularly with regard to residence properties and disease resistance. As a result, overall productivity of the carp in these systems can be superior to that of faster-growing, genetically improved varieties of the species, due to their properties of adaptation.
Factors in genetic variation
Four processes resulting from domestication can impact levels of genetic variation and therefore alter phenotypic characteristics: unintentional selection, inbreeding, genetic drift, and hybrid introgression.
Unintentional selection
Directional selection occurs as a result of unintentionally biased broodstock replacement strategies. In the long term, biased selection of fish that contribute to the next generations will result in changes in the genetic and phenotypic characteristics of the populations. These changes may even arise as indirect consequences of intentional selection, and can be either positive or negative.
Selecting larger fish
For example, in seasonal spawners such as Indian major carp, larger fish are often selected as future broodfish on the basis that they are more fecund and may grow faster. In reality, the larger fish within an age class are usually the ones produced early in the breeding season. In the long term, this selection for older fish within each year class will result in selection for early maturation.
Other selection traits
Another common example would be farms that sell their largest fish and retain the less-valuable, smaller fish for future broodstock, unintentionally selecting for slower growth rates. Monks who harvested common carp from monastery ponds using seine nets, leaving the remaining fish to breed in the pond, practiced unintentional selection for net avoidance. This is why the domesticated common carp is today one of the hardest fish to net from a pond. Unintentional selection commonly affects such other traits as sexual maturation and spawning time.
Inbreeding and inbreeding depression
Inbreeding is the loss of genetic variance (heterozygosity) as a consequence of the mating of close relatives. Inbreeding has the opposite effect of maintaining or enhancing genetic variation, with generally deleterious consequences known as inbreeding depression. Inbreeding depression is manifested in general loss of fitness, lower growth rates, poorer survival, increased incidence of deformities, or loss of fecundity.
Inbreeding increases homozygosity across all loci, fixing some alleles while others are lost. However, inbreeding several lines – for example, by continued sibmating (brother-sister matings) – will not necessarily mean the same alleles will be fixed in each line. To some extent, crossing between inbred lines (out breeding) will restore genetic variance and negate the effects of inbreeding. This method is used intentionally in some crossbreeding programs.
Genetic drift
Genetic drift reflects the random changes in gene frequencies that result from “sampling” a population, as opposed to unintentional or deliberate selection, which bring about directional changes. Sampling that results in these changes can be natural (e.g., escapes, disease outbreaks) or manmade (transfers, culling). Genetic drift leads to loss or fixation of alleles (variants of individual genes) within populations.
Founder effects are major agents in genetic drift, and occur when new hatchery populations are established from existing populations. Often a relatively small proportion of the original population (i.e., the sum of genetic variation) is sampled to provide the basis for new populations.
Introgression

Genetic or hybrid introgression is the mixing of genes between species or strains. This results initially from deliberate or accidental hybridization in hatcheries. If these hybrids are then used as broodstock, there is segregation of the genes, with the result that some of the fish carrying the introgressed genes cannot be easily distinguished from the original pure species. Thus, populations that contain a proportion of genes from another species develop.
Introgression is common in tilapia and other interbreeding species groups where hybrids can be easily produced artificially or naturally. Hybrid introgression is thought to have occurred in some Indian major carp populations where hybrids were produced, either out of scientific interest or through broodstock shortages. This introgression will likely have negative consequences from the loss of the pure species’ distinct feeding strategies, which are the basis for successful carp polyculture systems.
Effective population size
The ideal population size is infinitely large, ensuring no loss of genetic variation. In reality, hatchery managers must work with small, finite populations. The total number of broodfish held at a farm is not the most important factor of broodstock management, but rather the number that contribute genes to subsequent generations.
This effective population size (Ne) is a function of the total number of breeding individuals, sex ratio, mating system employed, and variance of family size in the production of fish for the next generation. In a random breeding population, Ne can be calculated from the following formula:
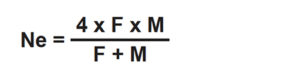
Ne should be maximized to minimize the loss of genetic variation. This can be achieved by retaining some progeny for future use as broodstock (preferably in equal numbers from each spawning) with a 1:1 ratio of contributing males and females.
When a population passes through a generation with low Ne, subsequent increases in Ne cannot recreate genetic variation lost as a result of inbreeding and genetic drift, but will limit any further loss of variation. When periodic reductions in Ne occur, genetic variation is irrevocably lost from the population with each bottleneck through which it passes.
What is the minimum Ne? Recommendations vary from 50 to 1,000, but inbreeding will rapidly accumulate, with deleterious implications if Ne is regularly below 50, as is common in many hatcheries.
Value of genetic variation
In domesticated populations of fish, it is desirable to maintain high levels of genetic variation for the following reasons:
- Genetic variation provides the raw material for selection. Without additive genetic variance, there can be no response to selection.
- Rare alleles in a population may not be of value to aquaculture today, but may be tomorrow (e.g., in conferring resistance to an outbreak of a new disease).
- Once alleles are lost from a population, they cannot be recovered without the addition of new material.
- Genetically variable populations are thought to have higher levels of developmental stability and fitness.
- Inbreeding depression is prevented.
Good Practices in broodstock management
- Avoid using hybrid fish as broodstock unless this is part of a planned breeding program.
- Spawn fish over the entire breeding season.
- Spawn fish of all sizes where possible, using broodstock of known age classes.
- Spawn as many fish as practically feasible.
- Spawn fish in equal ratios of males and females where possible.
- Maintain even family sizes up to the production of the next generation, to ensure that each mating pair or breeding set contributes genes to the next generation in roughly equal proportions.
- Tag broodstock if possible and keep full records on broodstock replacement practices.
Conclusion
The responsibility of hatchery managers for the maintenance of genetic quality of their stocks depends on the structure of the particular industry. For example, if a hatchery regularly receives future broodstock from a central broodstock supply center, the hatchery has minimal responsibility. However, where a hatchery population functions in reproductive isolation with no introductions, steps must be taken to prevent the harmful effects of domestication and maintain genetic variation within the population.
Research on the application of genetics in aquaculture in developing countries is increasingly focusing on broodstock management issues. Undoubtedly, there is much to learn from the mistakes that have been made to date in the domestication and management of a number of commercially important fish species. Improvements in basic management practices could have substantial long-term, positive impacts on aquaculture production.
(Editor’s Note: This article was originally published in the February 2002 print edition of the Global Aquaculture Advocate.)
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Author
-

School of Biological Sciences
University of Wales – Swansea, U.K.
Aquaculture & Aquatic Resources Management
Asian Institute of Technology
P.O. Box 4, Klong Luang
Pathumthani 12120 Thailand
Related Posts
Health & Welfare
Domestication of gut microbiota can improve shrimp aquaculture
In studies, bacteria and yeasts isolated from the native microbiota of wild shrimp at larval, juvenile and adult life stages were administered to early-stage shrimp via the culture water.

Health & Welfare
Black tiger domestication, selective breeding advance in Australia
Using clear-water tank systems, CSIRO and a collaborating farm have advanced the domestication of black tiger stocks in Australia.

Health & Welfare
Black tiger shrimp domestication advances
The production of early generations of domesticated broodstock in open-environment ponds may have hampered the domestication of black tiger shrimp.

Health & Welfare
Atlantic cod genomics and broodstock development project
The Atlantic Cod Genomics and Broodstock Development Project has expanded the gene-related resources for the species in Canada.